One of the toughest nuts to crack in population genetics has proved to be the story of the people of the Hindu Kush. However, using TreeMix and ancient genomes from the recent Allentoft et al. and Haak et al. papers, I'm seeing most of the Kalash and Pathan individuals from the HGDP modeled as ~65% Late Neolithic/Early Bronze Age (LN/EBA) European and ~35% Central Asian. This, to me at least, makes a lot of sense. For instance:
The Kalash and Pathan samples that can't be modeled in this way, at least with the reference populations that I'm using, are fitted within a framework that closely resembles the old two-way Ancestral South Indian/Ancestral North Indian model (ASI/ANI). They usually score ~12% admixture from the branch leading to the Dai of southern China, which is obviously the proxy for ASI.
Both of these models are correct; they just show the same thing in different ways. So if we mesh them together the Kalash and Pathans come out ~65% LNE/EBA European (which includes substantial Caucasus or Caucasus-related ancestry), ~12% ASI, and ~23% something as yet undefined.
If I had to guess, I'd say the mystery ~23% was Neolithic admixture from what is now Iran. But ancient DNA has thrown plenty of curve balls at us already, so that's a low confidence prediction, even though it does make good sense.
It's also interesting to see the migration edges running from the Ulchi of east Siberia to the LN/EBA Europeans. This might be a signal of minor Eastern non-African (ENA), in other words East Eurasian, admixture. Then again, it might just be the algorithm trying to compensate for something, like excess Eastern Hunter-Gatherer (EHG) ancestry.
The full output from my analysis can be downloaded here. The reference samples and markers are listed here and here.
See also...
The Poltavka outlier
The real thing
The enigma of the Kalash
From the Eneolithic to the Late Bronze Age vast areas of Eurasia were inhabited by a series of highly mobile and innovative groups that mostly relied on pastoralism for subsistence and, judging by their warlike grave goods, didn't mind a bit of biffo.
In Europe, where they first appeared, their archeological remains are generally classified as part of the Corded Ware Culture (or its Battle-Axe and Single Grave offshoots), and in Asia, where they expanded rapidly from the Trans-Urals to the Pamirs and south Siberia, as part of the Sintashta, Petrovka and Andronovo cultures.
It's likely that these groups had a profound impact on the Bronze Age world, including on Mycenaean Greece and Hittite Anatolia. The Sintashta Culture, for instance, is credited with the development of the spoked-wheel chariot, which became widely used in warfare all the way from Egypt to China.
Unfortunately, a lot of nonsense has been written on this topic in the past. In my view, one of the most sensible and up do date sources currently online is the thesis The Origin and Spread of the War Chariot by Elias Manuel Morgado Pinheiro.
Indeed, the obvious awesomeness of these ancient people has stirred much controversy about their origins and legacy. The academic consensus is that they were closely related, and that at least some of their ancestors were early Indo-Europeans from Eneolithic Eastern Europe. But a few archeologists have argued that the Corded Ware Culture was native to Central Europe, and others that the Sintashta population arrived in the Trans-Urals from Iran or even Syria.
Moreover, linguists generally consider the Sintashta/Andronovo people as the most likely candidates for the Proto-Indo-Iranians, and thus the precursors of the Indo-Aryans. But this is contested by many Indologists, who prefer to see the deepest roots of the Indo-Iranians closer to India and often oppose the idea of an Aryan conquest of South Asia during the Bronze Age.
In the near future, probably within the next couple of years, ancient genomics will leave very little room for debate in these matters and the arguments will cease, at least in mainstream academia.
But we already have a reasonable collection of ancient DNA from the relevant archeological cultures. Does it back the general consensus? Let's take a look, starting with the Y-chromosome data sorted by culture. The bracketed numbers are the sample sources, which are listed at the bottom of the post.
Corded Ware, Germany, Individuals 2,3,4 [1], R1a
Corded Ware, Germany, I0104 [3], R1a
Corded Ware, Germany, RISE434 [4], R1a
Corded Ware, Germany, RISE436 [4], R1a
Corded Ware, Poland, RISE1 [4], R1b?
Corded Ware, Germany, RISE446 [4], R1a
Corded Ware, Poland, RISE431 [4], R1a
Single Grave?, Denmark, RISE61 [4], R1a
Battle-Axe, Sweden, RISE94 [4], R1a
Battle-Axe?, Sweden, RISE98 [4], R1b
Sintashta, Trans-Urals, Russia, RISE386 [4], R1a
Sintashta, Trans-Urals, Russia, RISE392 [4], R1a
Andronovo, South Siberia, Russia, S07 [2], C
Andronovo, South Siberia, Russia, S10 [2], R1a
Andronovo, South Siberia, Russia, S16 [2], R1a
Andronovo, Altai region, Russia, RISE512 [4], R1a
Fascinating stuff. Keep in mind also that at higher resolution, most, if not all, of these R1a lineages are actually R1a1a1, which is estimated to be only around 5,000-6,000 years old based on full Y-Chromosome sequences. In other words, these groups were certainly closely related, and in large part the descendants of a patriarch who lived no earlier than the Middle or even Late Neolithic.
Now, based on that list it might seem as if both R1a and Corded Ware were indeed native to North-Central Europe. But this is not so.
R1a appears to be an Eastern Hunter-Gatherer (EHG) marker that in all likelihood failed to penetrate west of present-day Ukraine until the Late Neolithic, because it's missing in all the relevant samples before this period. So it probably first arrived in Central Europe with the Corded Ware people. We know that the Corded Ware people were foreign to Central Europe because their genome-wide genetic structure is starkly different from that of the Middle Neolithic farmers who lived there before them.
This is easy to demonstrate. The Principal Component Analyses (PCA) below show where two ancient samples cluster alongside a variety of present-day West Eurasians from the Human Origins dataset [3]. Note that Esperstedt_MN, a Middle Neolithic sample from a Baalberge Group burial in east-central Germany [3], looks more at home in Sardinia than Central Europe. On the other hand, the Corded Ware sample [4], also from east-central Germany, is sitting at the other end of the plot, among groups from the Volga-Ural region.
I'll throw in a few more PCA featuring Corded Ware, Battle-Axe, Sintashta and Andronovo genomes that offered enough data to be placed on the plots with a high degree of accuracy [4]. Note that the only clear outlier is RISE512, an Andronovo sample with an inflated level of East Eurasian admixture. If you're having trouble finding the ancient samples, download the PDF files and use the PDF search field.
Corded_Ware_RISE00_Estonia
Corded_Ware_RISE431_Poland
Battle-Axe_RISE94_Sweden
Battle-Axe_RISE98_Sweden
Single_Grave_RISE61_Denmark
Sintashta_RISE386_Russia
Sintashta_RISE392_Russia
Sintashta_RISE394_Russia
Sintashta_RISE395_Russia
Andronovo_RISE500_Russia
Andronovo_RISE503_Russia
Andronovo_RISE505_Russia
Andronovo_RISE512_Russia
However, the meat and potatoes of ancient genomics are formal statistics. So in part 2 of this series I'll explore the genetic ancestry and legacy of the so called badasses of the Bronze Age using the ADMIXTOOLS software package.
Update 28/04/2016: I'm gearing up to finally write the second part of the Badasses of the Bronze Age. But before I do that, I need to update the first part, because I now have two more sets of ancient samples that can be described as truly badass in the context of this series: Potapovka and Srubnaya.
Incredibly, seven out of the eight Potapovka and Srubnaya males belong to R1a. One of the Potapovka individuals turned out P1, but that was only because of a lack of data, otherwise, in all likelihood, he'd also belong to R1a.
Potapovka, Samara, Russia, I0419 [5], R1a
Srubnaya, Samara, Russia, I0232 [5], R1a
Srubnaya, Samara, Russia, I0360 [5], R1a
Srubnaya, Samara, Russia, I0361 [5], R1a
Srubnaya, Samara, Russia, I0423 [5], R1a
Srubnaya, Samara, Russia, I0424 [5], R1a
Srubnaya, Samara, Russia, I0430 [5], R1a
Also, I recently changed the way I run PCA. Indeed, I can now accurately plot samples on a typical genetic map of West Eurasia with as little as 15K SNPs. So instead of a few plots featuring the best Potapovka and Srubnaya sequences separately, here's a plot with all of the badasses together.
Update 22/09/2017: Unfortunately, I won't be able to finish this series anytime soon. Things are moving very quickly, but the full picture won't be known to us for a while yet. Some recent developments are covered at my other blog under these links...
R1a-M417 from Eneolithic Ukraine!!!11
Late PIE ground zero now obvious; location of PIE homeland still uncertain, but...
Citations...
1. Haak et al., Ancient DNA, Strontium isotopes, and osteological analyses shed light on social and kinship organization of the Later Stone Age, PNAS, Published online before print November 17, 2008, doi:10.1073/pnas.0807592105
2. Keyser et al., Ancient DNA provides new insights into the history of south Siberian Kurgan people, Human Genetics, Saturday, May 16, 2009, doi: 10.1007/s00439-009-0683-0
3. Haak et al., Massive migration from the steppe is a source for Indo-European languages in Europe, bioRxiv, Posted February 10, 2015, doi: https://dx.doi.org/10.1101/013433
4. Allentoft et al., Bronze Age population dynamics, selection, and the formation of Eurasian genetic structure, Nature 522, 167–172 (11 June 2015) doi:10.1038/nature14507
5. Mathieson et al., Genome-wide patterns of selection in 230 ancient Eurasians, Nature, 528, 499–503 (24 December 2015), doi:10.1038/nature16152
Ancient DNA tests on a skeleton from an Early Bronze Age "warrior" grave near Hrubieszow, southeastern Poland, have revealed that the remains belong to Y-haplogroup R1a1a [source].
Mitochondrial sequences were also obtained from seven other samples from the same burial site, and assigned to mt-haplogroups H1a, H1b (two), H2a (two), H6 and U5b1.
R1a1a is by far the most frequent Y-haplogroup in Poland today, and its presence in the remains from a high-status burial might be a clue as to how it became so common in East-Central Europe.
Interestingly, the site is classified as part of the Strzyżow Culture, which is considered by Polish archaeologists to be the result of contacts between local communities in southeastern Poland and Kurgan newcomers from the North Pontic steppe.
All of the other ancient R1a1a samples reported to date from Central Europe are also younger than the Middle Neolithic and from presumably steppe-derived Indo-European archeological cultures:
- Late Neolithic, Eulau, Germany, Corded Ware Culture, three related samples
- Late Neolithic, Esperstedt, Gemany, Corded Ware Culture, one sample
- Late Bronze Age, Halberstadt, Germany, Urnfield Culture (?), one sample
- Late Bronze Age, Lichtenstein Cave, Germany, Urnfield Culture, two samples
More info about the Bronze Age Pole, including photos of a facial reconstruction, can be found here and here (in Polish).
See also...
R1a-Z280 from Early Bronze Age Northern Poland
Testing for genetic continuity in Poland from the Bronze Age to the present
A new paper at PLoS ONE featuring ancient mitochondrial (mtDNA) data from Wielbark, Przeworsk and early Slavic remains argues for matrilineal continuity in present-day Poland since the Iron Age. It's actually based on a thesis that I blogged about more than two years ago (see here). However, it does include some fresh insights, so it's worth a look even if you read the thesis. RoIA stands for Roman Iron Age.
Three modern populations or groups of populations (Lithuanians and Latvians, Poles, and Czechs and Slovaks) were found to contain significantly higher percentages (p,0.05) of shared informative haplotypes with the RoIA samples compared to other present-day populations (Figure 2, Table S4). Notably, modern Poles shared the highest number (nine) of informative mtDNA haplotypes with the RoIA individuals.
...
Of particular interest are three RoIA samples assigned to subhaplogroup H5a1, which were recovered from the Kowalewko (sample K1), the Gaski, and the Rogowo (samples G1 and R3) burial sites (see Figure 1). Recent studies on mtDNA hg H5 have revealed that phylogenetically older subbranches, H5a3, H5a4 and H5e, are observed primarily in modern populations from southern Europe, while the younger ones, including H5a1 that was found among RoIA individuals in our study, date to around 4.000 years ago (kya) and are found predominantly among Slavic populations of Central and East Europe, including contemporary Poles [15]. Notably, we also found one ME sample belonging to subhaplogroup H5a1 (sample OL1 in Table 3). The presence of subclusters of H5a1 in four ancient samples belonging to both the RoIA and the ME periods, and in contemporary Poles, indicates the genetic continuity of this maternal lineage in the territory of modern-day Poland from at least Roman Iron Age i.e., 2 kya.
...
The evolutionary age of H5 sub-branches (,4 kya) [15] also approximates the age of N1a1a2 subclade found in the RoIA population (sample KA2) (Table 2). The coalescence age of N1a1a2 is around 3.4–4 kya, making this haplotype one of the youngest sub-branches within hg N [52]. The N1a1a2 haplotype found in one RoIA individual was classified as unique because no exact match was found among the twelve comparative populations or groups of populations used in the haplotype sharing test. Notably, a similar N1a1a2 haplotype carrying an additional transition at position 16172 was found in a modern-day Polish individual [53].

I suspect the publication of these results at this time, so many months after they were first revealed in the aforementioned thesis, is part of an effort to drum up interest and secure funding for a new project on the genetic history of Greater Poland, which was announced late last year (see here). I say that because one of the people organizing the project, Janusz Piontek, is also listed as a co-author on this paper. So if we're lucky we might soon see full genome sequences from a few of these Iron Age and Medieval samples.
Citation...
Juras A, Dabert M, Kushniarevich A, Malmstro¨m H, Raghavan M, et al. (2014) Ancient DNA Reveals Matrilineal Continuity in Present-Day Poland over the Last Two Millennia. PLoS ONE 9(10): e110839. doi:10.1371/journal.pone.0110839
A new paper in the Economics & Human Biology journal argues that male height in Europe is mostly determined by nutrition and genetics. That's not exactly earth shattering news. However, the authors also point out that Y-chromosome haplogroup I-M170 shows a strong correlation with the highest average stature on the continent, and speculate that the link between the two might be Upper Paleolithic hunter-gatherer ancestry:
The average height of 45 national samples used in our study was 178.3 cm (median 178.5 cm). The average of 42 European countries was 178.3 cm (median 178.4 cm). When weighted by population size, the average height of a young European male can be estimated at 177.6 cm. The geographical comparison of European samples (Fig. 1) shows that above average stature (178+ cm) is typical for Northern/Central Europe and the Western Balkans (the area of the Dinaric Alps). This agrees with observations of 20th century anthropologists (Coon, 1939; Lundman 1977). At present, the tallest nation in Europe (and also in the world) are the Dutch (average male height 183.8 cm), followed by Montenegrins (183.2 cm) and possibly Bosnians (182.5 cm) (Table 1). In contrast with these high values, the shortest men in Europe can be found in Turkey (173.6 cm), Portugal (173.9 cm), Cyprus (174.6 cm) and in economically underdeveloped nations of the Balkans and former Soviet Union (mainly Albania, Moldova, and the Caucasian republics).
...
The trend of increasing height has already stopped in Norway, Denmark, the Netherlands, Slovakia and Germany. In Norway, military statistics date its cessation to late 1980s.
...
In contrast, the fastest pace of the height increase (≥1 cm/decade) can be observed in Ireland, Portugal, Spain, Latvia, Belarus, Poland, Bosnia and Herzegovina, Croatia, Greece, Turkey and at least in the southern parts of Italy.
...
Although the documented differences in male stature in European nations can largely be explained by nutrition and other exogenous factors, it is remarkable that the picture in Fig. 1 strikingly resembles the distribution of Y haplogroup I-M170 (Fig. 10a). Apart from a regional anomaly in Sardinia (sub-branch I2a1a-M26), this male genetic lineage has two frequency peaks, from which one is located in Scandinavia and northern Germany (I1-M253 and I2a2-M436), and the second one in the Dinaric Alps in Bosnia and Herzegovina (I2a1b-M423)16. In other words, these are exactly the regions that are characterized by unusual tallness. The correlation between the frequency of I-M170 and male height in 43 European countries (including USA) is indeed highly statistically significant (r = 0.65; p < 0.001) (Fig. 11a, Table 4). Furthermore, frequencies of Paleolithic Y haplogroups in Northeastern Europe are improbably low, being distorted by the genetic drift of N1c-M46, a paternal marker of Ugrofinian hunter-gatherers. After the exclusion of N1c-M46 from the genetic profile of the Baltic states and Finland, the r-value would further slightly rise to 0.67 (p < 0.001). These relationships strongly suggest that extraordinary predispositions for tallness were already present in the Upper Paleolithic groups that had once brought this lineage from the Near East to Europe.

Citation...
Grasgruber et al., The role of nutrition and genetics as key determinants of the positive height trend, Economics & Human Biology, available online 7 August 2014, DOI: 10.1016/j.ehb.2014.07.002
There's been a lot of horseshit published over the years about Y-chromosome haplogroup R1a, which just happens to be my haplogroup. This includes academic papers in journals like PLoS ONE and Nature.
Indeed, a new paper on the phylogeography of R1a appeared at the Nature website today: Underhill et al. 2014. It's actually a much better effort than anything else on the topic at academic level thus far, but certainly not without issues.
For instance, the authors failed to include two well known and very important R1a subclades in their analysis: the Northwest European-specific R1a-CTS4385 and the East and Central European-specific R1a-Z280. As a result, the former is lumped with R1a-M417* and the latter with R1a-Z282*. In fact, Z280 is shown to be above Z282 in the topology of R1a-M420 (see Figure 1 here), which is plain wrong. These are major oversights and mean that this study is not a very useful resource as far as the phylogeography of European R1a is concerned.
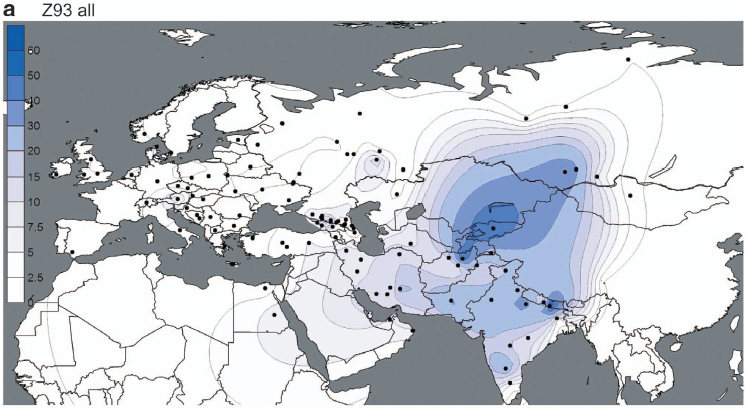
But the paper does show a couple of interesting things. For instance, the maps below offer the best illustration to date of the dichotomy between the European-specific R1a-Z282 and Asian-specific R1a-Z93.
However, these are very closely related subclades, sharing the Z645 mutation (unfortunately not mentioned in the paper), and both reaching high frequencies among Indo-European speakers. It's therefore plausible that groups carrying these markers expanded to the west and east from a zone between their current hotspots, possibly the Volga-Ural region, rather recently.
These migrations had to have happened after 4800-6800 YBP, which is the age of R1a-M417 reported by Underhill et al., and backed up by estimates from genetic genealogists using, among other things, complete R1a sequences (see here). In other words, the rapid expansions of R1a-Z282 and R1a-Z93 appear to have taken place from more or less the same region during the generally accepted early Indo-European timeframe, making them excellent candidates for paternal markers of the early Indo-European dispersals.
At the same time, the paucity of R1a-Z93 and derived lineages in Europe, including Eastern Europe, suggests that historic migrations originating in East and Central Asia, like those of the early Turks, had a negligible effect on the paternal ancestry of modern Europeans. This shows very clearly on the PCA in Figure 4 (see here).
Citation...
Underhill et al., The phylogenetic and geographic structure of Y-chromosome haplogroup R1a, European Journal of Human Genetics, advance online publication, 26 March 2014; doi:10.1038/ejhg.2014.50
See also...
The beast among Y-haplogroups
Over at Nature Communications, Rootsi et al. report on a newly discovered Ashkenazi-specific subclade of R1a, defined by the M582 mutation. They argue that it's a marker of Near Eastern origin, and based on the comprehensive data in their paper, I'd say they're correct. However, it's important to note that this doesn't preclude an ultimate Eastern European or Central Asian source of M582 in the Near East. For instance, an R1a mutation ancestral to M582 might have been introduced by the proto-Iranians from the steppe into what is now Iran during the early Indo-European dispersals. Indeed, that's actually what Figure 1a from the study suggests (phylogenetic tree of R1a below). The paper is open access, but here are a few quotes anyway:
Haplogroup R1a-M582 was only sporadically observed in Europe, the Diaspora residence of Ashkenazi Jews. Notably, it was not identified among 2,149 samples (including 922 R1a-M198) of non-Jews from East Europe, where the Ashkenazi Jewish community flourished in recent centuries (Table 1).
...
Within 1,068 West/North European samples (106 R1a-M198), M582 was observed in just one German sample, and among 3,756 Central/South European samples (710 R1a-M198), it was found only in one Hungarian and one Slovakian sample (Table 1).
...
Among 3,739 Near Eastern samples (303 R1a-M198), R1a-M582 was identified in various populations, with the highest frequency occurring within Iranians collected from the southeastern Kerman population who self-identified as Persians, northwestern Iranian Azeri and in Cilician Anatolian Kurds, at 2.86%, 2.50% and 2.83%, respectively (Table 1). In contrast, among 2,164 samples from the Caucasus (211 R1a-M198), R1a-M582 was found in just one Nogay sample (Table 1).
...
Considering the historical records of Ashkenazi Jews, three potential geographic sources should be considered: the Near East, which was the geographic location for the ancient Hebrews; Europe, which was the residence of the Ashkenazi Jewish Diaspora and the region in which they evolved for nearly two millennia; and the region overlapping with the no longer extant mid-11th Century Khazarian Khaganate, whose ruling class has been suggested to have converted to Judaism [18]. Our data render the latter source highly unlikely since the Khazarian Khaganate overlapped with the Northern Pontic-Caspian steppe and the North Caucasus region, in which just one Nogay sample carried the R1a-M582 haplogroup (Table 1). Furthermore, the Nogays, formerly a powerful Kipchak Turkic-speaking nomadic confederation, are relatively recent inhabitants of the Caucasus, and the STR haplotype of the sole R1a-M582 Nogay sample lies outside of the Levite cluster. Had the Caucasus region been the source for the Ashkenazi modal lineage, we likely would have found R1a-M582 Y-chromosomes in some of its 20 local populations examined in our sample of more than 2,000 Y-chromosomes (Table 1).
...
Near Eastern populations are the only populations in which haplogroup R1a-M582 was found at significant frequencies (Table 1). Moreover, the representative samples displayed substantial diversity even within this geographic region (Fig. 1b). Higher frequencies and diversities often suggest lineage autochthony.

Citation...
Rootsi, S. et al. Phylogenetic applications of whole Y-chromosome sequences and the Near Eastern origin of Ashkenazi Levites. Nat. Commun. 4:2928 doi: 10.1038/ncomms3928 (2013).
See also...
The Poltavka outlier
He was short (<160 cm), probably lactose intolerant, had an exceedingly long melon (cranial index = 72.6), and belonged to mtDNA haplogroup K2a. In other words, he was a typical Neolithic farmer, and clearly different from the average modern-day inhabitant of the North European Plain.
No doubt, his people were largely replaced by newcomers from the east and also west during the frequent population shifts in the region after the Neolithic (see here). However, the stable isotope analysis suggests that he ate a lot of millet, which is known as a typically Slavic cultigen in Europe.
ABSTRACT: In 2007 a ceremonial complex representing the Globular Amphora Culture was discovered in Kowal (the Kuyavia region, Poland). Radiocarbon dating demonstrated that the human remains associated with the complex are of similar antiquity, i.e. 4.105 ± 0.035 conv. and 3.990 ± 0.050 conv. Kyrs. After calibration, this suggests a period between 2850 and 2570 BC (68.2% likelihood), or more specifically, 2870 to 2500 BC (95.4% likelihood). Morphological data indicate that the skeleton belonged to a male who died at 27–35 years of age. The unusual morphology of his hard palate suggests this individual may have had a speech disorder. Stable oxygen isotope values of the individual's teeth are above the locally established oxygen isotope range of precipitation, but due to sample limitations we cannot conclusively say whether the individual is of non-local origin. Stable carbon and nitrogen isotope ratios were analyzed to reconstruct the diet of the studied individual, and show a terrestrial-based diet. Through ancient DNA (aDNA) analysis, the mtDNA haplogroup K2a* and lactose intolerance as evidenced by homozygous C-13910 allele were identified. These aDNA results are the first sequences reported for an individual representing the Globular Amphora Culture, enriching the still modest pool of human genetic data from the Neolithic.

Kozłowski T., Stepańczak B., Laurie J. Reitsama, Osipowicz G., Szostek K., Płoszaj T., Jędryhowska-Dańska K., Pawlyta J., Paluszkiewicz C., Witas H.W. Osteological, chemical and genetic analyses of the human skeleton from a Neolithic site representing the Globular Amphora Culture (Kowal, Kuyavia region, Poland), Anthropologie [In Press]
See also...
Polish "Goths" enjoyed their millet, while Polish "Vikings" did not
One of the most enduring myths or cliches concerning European genetic structure is that Poles carry Mongolian admixture. This claim has been repeated so often that it's now regarded by many as fact, including at academic level. Poles apparently acquired this admixture during Mongol and Turkic raids on Eastern Europe during the Middle Ages.
For a long time it was impossible to verify or debunk such claims due to a lack of genetic data from European and Asian populations at high enough resolution. However, that's no longer a problem. Indeed, I've run a wide range of detailed analyses as part of my Eurogenes Genetic Ancestry Project which have shown that my Polish sample does not carry inflated levels of Asian ancestry relative to other Northern and Central Europeans. But in this post it's probably more useful for me to focus on results from peer-reviewed studies to make my point.
I'll start with a look at genome-wide genetic data (aka. autosomal DNA). The bar plot below was featured in a recent study on genetic substructures in European Russia (1). It shows the results of an ADMIXTURE analysis at K=2 (two ancestral populations assumed) using 52,808 SNP markers, which splits the genomes of the samples into West and East Eurasian components. As you'd expect, these components are modal in Europeans and Han Chinese, respectively. This northern Han Chinese sample from Beijing is certainly a very useful proxy for Mongolians, because the East Eurasian component it creates is basically the same one that peaks in the least admixed Mongolian samples in other studies (for example, see 2).
The lowest levels of the East Eurasian component are carried by Latvians, Poles, Germans, Czechs and Italians. But this looks like noise anyway, because the Chinese carry a reciprocal amount of the West Eurasian component. In other words, it's most likely not a real signal of recent admixture from East Asia, but shared prehistoric Eurasian ancestry. If it's not noise, and actually represents admixture from an East Asian source like the Mongolians, then it's difficult to explain why it appears at a clearly higher level in Italians than Poles.
A couple of the Czechs do carry inflated amounts of the East Eurasian component, and I'd say they're Czech Roma with significant South Asian admixture who weren't removed as outliers from the dataset. It's also worth mentioning that the Komi, Finns and Russians (like the Rus_HGDP sample from Kargopol) show higher levels of this component than the Central Europeans. However, this doesn't necessarily mean they have Mongolian ancestry. Indeed, another study has shown that the Kargopol Russians carry various Siberian-specific components, rather than the type of East Asian influence which makes up the majority of Mongolian genome-wide genetic structure (2).
Moreover, the Mongols never raided North Russia or Finland, where in fact these components peak in Europe today. So the most plausible explanation for the relatively high levels of Siberian admixture there is Finno-Ugric or Uralic ancestry. Indeed, the Finns obviously speak a Finno-Ugric language, and so do some North Russian groups, while many others did until recently.
Below is a different kind of analysis of autosomal DNA. It's a pairwise Fixation Index (Fst) test between 16 global populations based on 129,673 SNPs (3). It includes two East Asian samples, the same Han Chinese set as in the above ADMIXTURE analyses, and a Japanese sample from Tokyo. The results appear to correlate very closely with geography, which suggests that we're basically looking at the effects of isolation-by-distance. Interestingly, the Polish sample from Lodz and Warsaw (Po) is genetically more distant from the Han Chinese (CHB) and Japanese (JPT) than are the Swedes (Sw), Norwegians (No) and Germans (Ge). The differences aren't big, but they're consistent, which means that at the very least these Poles can't be carrying more East Asian ancestry than the Scandinavian and German samples.
Next up is a global Principal Component Analysis (PCA) from the same study, featuring the same samples and markers. Again, it's another way of looking at variation in autosomal DNA, and again the results indicate that Poles don't show any special genome-wide genetic links to East Asians. The Polish samples are sitting close to the top of the European cluster and overlap strongly with other Northern and Central Europeans. Indeed, many of the outliers pulling towards the West African (YRI) and East Asian (CHB + JPT) clusters are French, British and German. These individuals are possibly carrying very recent non-European ancestry due to colonial links between Western Europe and the third world. There are also a considerable number of Russians streaming towards the East Asians, and that's probably due to the Finno-Ugric mediated Siberian influence in North Russia described above.
 It might also be useful to assess the level of West Asian or Near Eastern autosomal admixture in Poland compared to other parts of Europe, because many of the Turkic groups which ended up on the Eastern European steppe were largely of West Asian origin. Therefore, if there was a significant genetic contribution from such groups to the Polish gene pool, then Poles today ought to show inflated affinity to West Asian populations compared to other Central and Northern Europeans. The simple answer is that they don't, which can be clearly seen on the pairwise Fst clustering analysis below based on approximately 101,000 SNPs (4).
Y-chromosome or paternal markers tell the same story as autosomal DNA. The most common Y-DNA haplogroup in Poland is R1a1a, making up about 50% of all the Y-chromosome lineages in the country. Based on academic and commercial testing of hundreds of Polish samples to date, it's safe to say that the most dominant subclades of this Eurasian haplogroup in Poland are the European-specific R-M458 and R-Z280 (5, 6). These subclades are closely related to the Scandinavian-specific R-Z284, because all three markers come off the Z283 branch of the R1a1a haplotree. The Z283 mutation is also mostly confined to Europe and possibly originated there.
The main Asian-specific subclade of R1a1a, known as R-Z94, appears to be extremely rare in Poland. This is the subclade that makes up the main share of the R1a1a in Mongolian and Turkic populations of Central Asia, as well as in South Asians. However, even the very few R-Z94 cases among Poles are more closely related to Ashkenazi lineages than those of Mongolians or Central Asians. What all of this means, of course, is that Mongolian or Turkic paternal ancestry isn't "hiding" in Poland under all that R1a1a.
The most common Y-DNA haplogroup among Mongolians and their close ethnic kin is C3. There's actually a particular lineage of C3 called the "star-cluster chromosome" which pops up regularly in purported descendents of the infamous Mongolian warlord Genghis Khan. This might mean that it's a marker of paternal ancestry from Genghis himself, which has been suggested in several academic papers (7, 8, 9). In any case, the haplotype is considered to be very young, probably less than 1000 years, and widespread in areas of Asia where the Mongol hordes were most active during the early Middle Ages. Therefore, it seems to be an excellent signal of Mongolian genetic influence from this key period. Below is a map of its frequencies in a variety of Asian groups.
It might also be useful to assess the level of West Asian or Near Eastern autosomal admixture in Poland compared to other parts of Europe, because many of the Turkic groups which ended up on the Eastern European steppe were largely of West Asian origin. Therefore, if there was a significant genetic contribution from such groups to the Polish gene pool, then Poles today ought to show inflated affinity to West Asian populations compared to other Central and Northern Europeans. The simple answer is that they don't, which can be clearly seen on the pairwise Fst clustering analysis below based on approximately 101,000 SNPs (4).
Y-chromosome or paternal markers tell the same story as autosomal DNA. The most common Y-DNA haplogroup in Poland is R1a1a, making up about 50% of all the Y-chromosome lineages in the country. Based on academic and commercial testing of hundreds of Polish samples to date, it's safe to say that the most dominant subclades of this Eurasian haplogroup in Poland are the European-specific R-M458 and R-Z280 (5, 6). These subclades are closely related to the Scandinavian-specific R-Z284, because all three markers come off the Z283 branch of the R1a1a haplotree. The Z283 mutation is also mostly confined to Europe and possibly originated there.
The main Asian-specific subclade of R1a1a, known as R-Z94, appears to be extremely rare in Poland. This is the subclade that makes up the main share of the R1a1a in Mongolian and Turkic populations of Central Asia, as well as in South Asians. However, even the very few R-Z94 cases among Poles are more closely related to Ashkenazi lineages than those of Mongolians or Central Asians. What all of this means, of course, is that Mongolian or Turkic paternal ancestry isn't "hiding" in Poland under all that R1a1a.
The most common Y-DNA haplogroup among Mongolians and their close ethnic kin is C3. There's actually a particular lineage of C3 called the "star-cluster chromosome" which pops up regularly in purported descendents of the infamous Mongolian warlord Genghis Khan. This might mean that it's a marker of paternal ancestry from Genghis himself, which has been suggested in several academic papers (7, 8, 9). In any case, the haplotype is considered to be very young, probably less than 1000 years, and widespread in areas of Asia where the Mongol hordes were most active during the early Middle Ages. Therefore, it seems to be an excellent signal of Mongolian genetic influence from this key period. Below is a map of its frequencies in a variety of Asian groups.
 However, this marker has never been recorded in Poland, where C3 itself is extremely unusual. Indeed, Derenko et al. conclude that based on the paucity of the "star-cluster haplotype" among ethnic Russians, the Mongol hordes of the Middle Ages didn't even leave a genetic imprint on European Russia.
However, this marker has never been recorded in Poland, where C3 itself is extremely unusual. Indeed, Derenko et al. conclude that based on the paucity of the "star-cluster haplotype" among ethnic Russians, the Mongol hordes of the Middle Ages didn't even leave a genetic imprint on European Russia.
It is known that the Mongol Empire expanded over a considerable part of Eastern Europe by 1248 due to the khan Batu’s conquests. Russian principalities were vassal states of the Mongol Empire until 1480. However, we found no genetic traces of the Mongol sovereignty over Russia (in the form of male lineages of the cluster of Genghis Khan descendants) in the Russian population.
Poles do sporadically carry East Eurasian-specific mtDNA (ie. maternal) lineages, and it was recently suggested by Mielnik-Sikorska et al. that some of these lineages (ie. C4a1a, G2a and D5a2a1a1) might "possibly reflect relatively recent contacts of Slavs with nomadic Altaic peoples" (10). However, the authors also note that these markers have a wide distribution across Eurasia and might actually represent prehistoric gene-flow between Asia and Europe.
In fact, ancient DNA results have recently revealed unexpected frequencies of East Eurasian-specific mtDNA haplogroups - including C1, C4a2, C5, D* and Z1a - in Northern and Eastern European remains from the Neolithic and Mesolithic (11, 12, 13, 14). The samples were always small, but nevertheless it's useful to note that the incidence of the eastern mtDNA lineages was much higher in these ancient samples than in any modern Eastern European populations west of the Volga. What this suggests is that the vast majority of East Eurasian ancestry in Europe might have arrived there thousands of years before the Mongol incursions, and much of it has been lost since then, rather than gained, due to continuous population movements from west to east across the continent.
To add another twist to the tale, it actually seems that all Europeans do show significant prehistoric East Eurasian genome-wide admixture, so perhaps it's only the ancient eastern mtDNA markers that have almost gone extinct? The topic has been now been discussed in a couple of papers, and apparently this admixture shows higher affinity to modern Amerindians than North or East Asians (15), which is very curious indeed. But the details are sketchy because the results so far have been based on DNA from modern samples, so hopefully we can learn more very soon thanks to analyses of genome-wide markers from prehistoric Eurasian remains. In any case, a group of Poles was featured in one of these papers, and this is how they compared to other Europeans in a formal mixture test with the ADMIXTOOLS software (16).
 The positions of the samples in the table are based on the "Sardinian" f3-statistic, which indicates the strength of the Amerindian-like admixture signal when Karitiana Indians and Sardinians are chosen as references. The results appear fairly random in some cases, and that's probably because they're skewed by such factors as genetic drift and sample size - for instance, heavy drift might dampen the signal, while a larger sample size might increase it. But the outcomes are also clearly influenced by relatively recent admixture from Siberia, Central Asia and/or the Indian subcontinent. That's because the samples known to carry such admixtures, like the HGDP Russians and Turks, are at the top of the table. Therefore, it's worth noting that the Polish sample is found at the bottom, which is in line with results from all the other analyses presented above.
Please note that the information in this post pertains to the current Polish population by and large, and to individuals with all grandparents of ethnic Polish origin. It's not relevant to individuals whose recent ancestors came from within the borders (or former borders) of Poland but their ethnic origins were uncertain. Keep in mind that Poland was not as ethnically or genetically homogenous before World War II as it is today. Also worth noting is that in a country of almost 40 million people, some individuals will have very atypical pedigrees, and it might even be possible to find someone with, say, Papuan ancestry in Poland if we look hard enough.
By the way, I've no idea who made that meme at the top of the post, or where it was published originally, but I think it's very appropriate here. Please let me know if it violates any copyright laws and I'll take it down.
References...
1. Khrunin AV, Khokhrin DV, Filippova IN, Esko T, Nelis M, et al. (2013) A Genome-Wide Analysis of Populations from European Russia Reveals a New Pole of Genetic Diversity in Northern Europe. PLoS ONE 8(3): e58552. doi:10.1371/journal.pone.0058552
2. Morten Rasmussen et al., Ancient human genome sequence of an extinct Palaeo-Eskimo, Nature 463, 757-762 (11 February 2010) doi:10.1038/nature08835; Received 30 November 2009; Accepted 18 January 2010
3. Simon C Heath et al, Investigation of the fine structure of European populations with applications to disease association studies, European Journal of Human Genetics (2008) 16, 1413–1429; doi:10.1038/ejhg.2008.210
The positions of the samples in the table are based on the "Sardinian" f3-statistic, which indicates the strength of the Amerindian-like admixture signal when Karitiana Indians and Sardinians are chosen as references. The results appear fairly random in some cases, and that's probably because they're skewed by such factors as genetic drift and sample size - for instance, heavy drift might dampen the signal, while a larger sample size might increase it. But the outcomes are also clearly influenced by relatively recent admixture from Siberia, Central Asia and/or the Indian subcontinent. That's because the samples known to carry such admixtures, like the HGDP Russians and Turks, are at the top of the table. Therefore, it's worth noting that the Polish sample is found at the bottom, which is in line with results from all the other analyses presented above.
Please note that the information in this post pertains to the current Polish population by and large, and to individuals with all grandparents of ethnic Polish origin. It's not relevant to individuals whose recent ancestors came from within the borders (or former borders) of Poland but their ethnic origins were uncertain. Keep in mind that Poland was not as ethnically or genetically homogenous before World War II as it is today. Also worth noting is that in a country of almost 40 million people, some individuals will have very atypical pedigrees, and it might even be possible to find someone with, say, Papuan ancestry in Poland if we look hard enough.
By the way, I've no idea who made that meme at the top of the post, or where it was published originally, but I think it's very appropriate here. Please let me know if it violates any copyright laws and I'll take it down.
References...
1. Khrunin AV, Khokhrin DV, Filippova IN, Esko T, Nelis M, et al. (2013) A Genome-Wide Analysis of Populations from European Russia Reveals a New Pole of Genetic Diversity in Northern Europe. PLoS ONE 8(3): e58552. doi:10.1371/journal.pone.0058552
2. Morten Rasmussen et al., Ancient human genome sequence of an extinct Palaeo-Eskimo, Nature 463, 757-762 (11 February 2010) doi:10.1038/nature08835; Received 30 November 2009; Accepted 18 January 2010
3. Simon C Heath et al, Investigation of the fine structure of European populations with applications to disease association studies, European Journal of Human Genetics (2008) 16, 1413–1429; doi:10.1038/ejhg.2008.210
4. Esko et al., Genetic characterization of northeastern Italian population isolates in the context of broader European genetic diversity, European Journal of Human Genetics advance online publication 19 December 2012; doi: 10.1038/ejhg.2012.229
5. Peter A Underhill et al., Separating the post-Glacial coancestry of European and Asian Y chromosomes within haplogroup R1a, European Journal of Human Genetics advance online publication 4 November 2009; doi: 10.1038/ejhg.2009.194
6. Family Tree DNA R1a1a and Subclades Y-DNA Project
7. Derenko et al., Distribution of the Male Lineages of Genghis Khan’s Descendants in Northern Eurasian Populations, Russian Journal of Genetics, 2007, Vol. 43, No. 3; DOI: 10.1134/S1022795407030179
8. Zerjal et al., The Genetic Legacy of the Mongols, Am J Hum Genet. 2003 March; 72(3): 717–721. PMCID: PMC1180246
9. Abilev S. et al., The Y-chromosome C3* star-cluster attributed to Genghis Khan's descendants is present at high frequency in the Kerey clan from Kazakhstan, Hum Biol. 2012 Feb;84(1):79-89. doi: 10.3378/027.084.0106.
10. Mielnik-Sikorska M, Daca P, Malyarchuk B, Derenko M, Skonieczna K, et al. (2013) The History of Slavs Inferred from Complete Mitochondrial Genome Sequences. PLoS ONE 8(1): e54360. doi:10.1371/journal.pone.0054360
11. Zsuzsanna Guba et al., HVS-I polymorphism screening of ancient human mitochondrial DNA provides evidence for N9a discontinuity and East Asian haplogroups in the Neolithic Hungary, Journal of Human Genetics advance online publication 15 September 2011; doi: 10.1038/jhg.2011.103
12. Alexey G Nikitin et al., Mitochondrial haplogroup C in ancient mitochondrial DNA from Ukraine extends the presence of East Eurasian genetic lineages in Neolithic Central and Eastern Europe, Journal of Human Genetics advance online publication, 7 June 2012; doi:10.1038/jhg.2012.69
13. Lillie, Malcolm C et al., Prehistoric populations of Ukraine: Migration at the later Mesolithic to Neolithic transition, Population Dynamics in Prehistory and Early History (2012), Publication Date: July 2012, ISBN: 978-3-11-026630-6, DOI: 10.1515/9783110266306.93
14. Der Sarkissian C, Balanovsky O, Brandt G, Khartanovich V, Buzhilova A, et al. (2013) Ancient DNA Reveals Prehistoric Gene-Flow from Siberia in the Complex Human Population History of North East Europe. PLoS Genet 9(2): e1003296. doi:10.1371/journal.pgen.1003296
15. Lipson et al., Efficient moment-based inference of admixture parameters and sources of gene flow, arXiv:1212.2555v1 [q-bio.PE]
16. Patterson et al., Ancient Admixture in Human History, Genetics: Published Articles Ahead of Print, published on September 7, 2012 as 10.1534/genetics.112.145037
See also...
R1a and R1b from an early Mongolian tomb
I've just been sent a fascinating thesis on the mtDNA of Iron Age and Medieval samples from Poland. It suggests direct genetic continuity between Iron Age samples belonging to the Przeworsk and Wielbark Cultures, of what is now West and Central Poland, and present-day Poles. Here's the English summary, and a map of the sites under study:
For many years the origin of the Slavs has been the subject-matter in archaeology, anthropology, history, linguistics and recently also modern human population genetics. By now there is no unambiguous answer to a question where, when and in what way the Slavs originated. For the purposes of this dissertation, the analysis of ancient human mitochondrial DNA was applied. The ancient DNA was isolated from 72 specimens which came from Iron-Age and medieval graveyards from the area of current Poland. Ancient mtDNA was extracted from two teeth from each individual and reproducible sequence results were obtained for 20 medieval and 23 Iron-Age specimens. On the basis of HVR I mtDNA mutation motifs and coding region SNPs each specimen was assigned to a mitochondrial haplogroup. The obtained results were used together with other ancient and modern populations to analyse shared haplotypes and population genetic distances illustrated by multidimentional scaling plots (MDS). The differences on genetic level and quite high genetic distances (FST) between medieval and Iron-Age populations as well as significant number of shared informative haplotypes with Belarus, Ukraine and Bulgaria may evidence genetic discontinuity between medieval and Iron Ages. From the other side, the highest number of shared informative haplotypes between Iron-Age and extant Polish population as well as the presence of subhaplogroup N1a1a2, can confirm that some genetic lines show continuity at least from Iron Age or even Neolithic in the areas of present day Poland. The results obtained in this work are considered to be the first ancient contribution in genetic history of the Slavs.

Below is an MDS from the thesis, based on data corrected for the effects of potential relatives in the Iron Age sample. I don't think it's a particularly useful way of judging the intra-European affinity of the two ancient Polish groups, mostly because the samples are small, and contemporary North, Central and East Europeans don't differ very much in terms of mtDNA. Nevertheless, we can see that both the Iron Age (Okres Rzymski) and Medieval (Sredniowiecze) samples fall within the range of modern European mtDNA diversity. On the other hand, the German Neolithic LBK sample (Neolit LBK Niemcy) clearly does not, because it's sitting at the far right of the plot, away from the main European cluster. This dichotomy between the genetic structure of the LBK farmers and modern Europeans has been demonstrated in previous studies, but the reasons for it are still a mystery.
Interestingly, modern Poles are closer to an Iron Age sample from Denmark (Okres Zelaza Dania) than to the Polish Iron Age set. However, as per the summary above, the author also compared the frequencies of the most informative haplotypes among the modern and ancient samples, and found that extant Poles are the closest group to the Polish Iron Age remains, followed by Balts, Swedes and Baltic Finns. Below is a table showing those results.
According to the author, these matches might hint at Baltic, Germanic and Finno-Ugric influences in the Polish Iron Age population. Perhaps, but in my opinion, they're simply in line with geography, and reflect the general North European character of maternal lineages shared by populations from around the Baltic, both today and during the Iron Age.
The results for the Medieval Polish sample are more intriguing, because they're somewhat out of whack with geography. Its best matching modern groups are Belorussians, Ukrainians and Bulgarians. This might suggest that, during the early middle ages, the territory of present day Poland experienced an influx of groups from what are now Belarus and Ukraine, who then melted into the gene pool of the natives of Polish Iron Age descent. However, conversely, it might mean that Belorussians, Ukrainians and Bulgarians descend in large part from fairly specific medieval groups from the area of modern Poland.
In any case, whether present day Polish territory saw some migrations from the immediate east during the Medieval period or not, this preliminary look at ancient Polish mtDNA suggests long-standing genetic continuity in the region. What it clearly doesn't show is a complete, or almost complete, population replacement in the areas between the Oder and Bug rivers during the migration period.
Indeed, the thesis results put into doubt past notions that the Przeworsk and Wielbark cultures were of Germanic origin.
The (mtDNA) haplogroup missing from both the Iron Age and medieval samples from the territory of modern Poland was haplogroup I. In contemporary Slavic populations, this haplogroup is found at levels ranging from 1.2% in Bulgarians to 4.8% in Slovaks. It was also recorded at high levels in ancient remains from Denmark. It showed a frequency of 12.5% in an Iron Age sample, and 13.8% in a medieval sample. Melchior et al. 2008 suggest that haplogroup I might have been more common in Denmark and Northern Europe during that period. Therefore, the lack of this haplogroup in ancient DNA from the territory of modern Poland, might mean that the Przeworsk and Wielbark cultures should not be identified with Germanic populations.
I'm sure more ancient DNA studies are on the way looking at the origins of Slavs and Poles. Indeed, if the Y-chromosomes of Przeworsk and Wielbark remains are successfully tested, I won't be surprised if they look fairly typical of modern Poles, with a decent representation of R1a1a-M458, which is the most common Y-chromosome haplogroup in Poland today.
Anna Juras, Etnogeneza Słowian w świetle badań kopalnego DNA, Praca doktorska wykonana w Zakładzie Biologii Ewolucyjnej Człowieka Instytutu Antropologii UAM w Poznaniu pod kierunkiem Prof. dr hab. Janusza Piontka